이번 포스트부터는 진핵생물 번역개시에 관여하는 각종 factor들에 대해 알아보자.

eukaryote에서의 translation initiation 과정을 간략히 나타낸 그림이 위와 같음.
우선 eIF3는 40S ribosomal subunit에 붙어 60S subunit과의 reassociation을 막아주고, 동시에 eIF6는 60S subunit에 붙어 40S subunit과의 reassociation을 막아줌. 한편 GTP가 붙은 상태의 eIF2는 Met-tRNA를 ribosome으로 데려오는 역할을 함. 이후 eIF2의 GTP가 hydrolysis되고 나면 eIF2는 40S subunit으로부터 release됨. 이 때 가수분해에 의해 생성된 GDP에는 (GEF[guanine nucleotide exchange factors]의 일종인) eIF2B가 작용해 이 녀석을 GTP로 다시 바꿔주게 됨. 이 상태에서 eIF4는 cap-binding protein으로 작용해 40S subunit이 mRNA의 5' end에 붙을 수 있도록 도와줌. 이후 eIF1과 eIF1A가 ribosome의 scanning을 도와주게 됨. 마지막으로 eIF5는 favorable한 initiation site 부근에서 48S complex와 60S ribosome subunit 간의 association을 매개해줌. 이런 과정을 거쳐 eukaryotic translation initiation이 이루어짐.
(참고로 bacteria에서의 IF2-GTP에서 GTP의 가수분해는 ribosome complex 전체가 있어야만 일어나는 event이기는 하나, 이 때 결과적으로 GTP를 가수분해시켜주는 녀석은 IF2임. 이 때의 activity를 ribosome-associated GTPase activity라고 부름. 이후 가수분해된 GDP는 ribosome 밖으로 release되게 됨)
function of eIF2

위 그림에는 eIF2-GTP에 의해 Met-tRNA가 recruit된 이후 eIF2에 붙어있는 GTP가 GDP로 바뀌고, 이에 의해 eIF2가 40S subunit으로부터 release되는 과정, 이후 GDP가 eIF2B에 의해 다시금 GTP로 바뀌는 과정이 나타나 있으므로 참고할 것.
function of eIF4

eIF4는 위 실험 결과에 나타나 있는 것처럼 3가지 polypeptide들로 이루어져 있음. 각각 eIF4E(25kD), eIF4A(50kD), eIF4G(220kD)이며 저마다의 역할이 존재함.

위 그림에 각각의 polypeptide들이 나타나 있음. 하나하나 살펴보자. 우선 eIF4E는 위 그림상에도 나와있는 것처럼 5' cap을 직접적으로 인식해 binding하는 역할을 수행함. 다음으로 eIF4G는 eIF4E와 연결되며 여타 다른 element들이 recruit될 수 있도록 해주는 scaffold 역할을 수행함. 마지막으로 eIF4A의 경우 helicase activity를 가져서 앞서 봤던 hairpin loop를 unwind시켜주는 역할을 수행함. 이 녀석이 있기에 적당한 높이의 hairpin이 존재하는 경우 그 hairpin이 풀리며 translation이 일어날 수 있는 것임. 이 때 eIF4A는 당연히 helicase activity를 가지므로 ATP dependent함.
한편 위 그림을 보면 eIF4G에는 eIF3와 Pab1p도 recruit되어 있다는 것을 알 수 있음. 이 때 eIF3는 원핵생물에서의 IF3와 마찬가지로 40S에 붙어 40S와 60S간의 reassociation을 막아주는 역할을 수행함. 한편 Pab1p는 mRNA 3' 말단의 poly A tail을 인지해서 결합함. 결과적으로 이들 complex에 의해 위와 같이 mRNA가 원형으로 감기게 됨. 이런 circular한 구조의 mRNA는 translation 조절, re-initiation의 촉진을 시켜줄 뿐 아니라, RNase로부터의 protection을 더 용이하게 만들어줌.
사실 이 밖에 eIF4를 이루는 accessory protein인 eIF4B도 존재함. 이 녀석은 RNA-binding domain을 가지고 있으며 eIF4A와 mRNA 간의 binding을 stimulate해주는 역할을 함. 이와 관련된 실험 결과가 아래에 나타나 있음.

이 실험에 사용한 것은 위 그림 오른쪽에 나온 40nt 길이의 nucleotide임. 보면 이 녀석들을 넣어줄 시 10bp 길이의 nucleotide 부분에 대해 상보적인 결합이 형성되면서 duplex가 형성됨. (각각의 sequence 5' 말단에는 radioactive한 인산기가 달려있음) 이러한 상황에서 eIF4A에 의한 helicase activity에 의해 이 duplex가 풀어져 monomer가 생성되는지 여부를 확인하기 위해 위 그림 왼쪽과 같은 gel mobility shift assay를 수행함.
lane 3과 5를 비교할 시 eIF4B가 없을 때에 비해 있을 때 monomer로의 분해가 더 잘 일어남을 확인함. 한편 lane 5, 6, 7을 비교할 시 점차 eIF4A를 많이 넣어줄수록 monomer가 되는 녀석의 비율이 더욱 증가함. 이를 통해 eIF4A가 helicase activity를 가지고 eIF4B는 eIF4A를 보조해주는 녀석이라는 것을 알 수 있음. 한편 lane 8과 같이 eIF4A를 넣어주지 않을 경우 eIF4B만 있을 경우에는 duplex의 분해가 일어나지 않았다는 것을 통해 eIF4B가 보조적인 역할을 한다는 것을 다시금 확인할 수 있음.
한편 virus 중 일부, 예를 들어 poliovirus와 같은 녀석들은 viral mRNA의 5' 부분에 capping이 일어나 있지 않아서 그 자체로는 host의 translation system을 이용하기 어려움. 이 때 이 녀석들이 사용하는 전략이 꽤나 흥미로우므로 이에 대해 잠시 알아보자.

poliovirus는 viral protease를 생산하는데, 이 viral protease는 위 그림상에도 나타나 있는 것처럼 eIF4G의 N-terminal domain을 잘라버림. 그렇게 되면 원래는 이 녀석과 함께 붙어야 할 eIF4E도 recruit되지 않을 것임. (즉, capping이 굳이 인식되지 않아도 complex가 형성될 것임) 이와 동시에 poliovirus는 mRNA 내에 internal ribosome entry sequence(IRES)라는 것을 가지고 있고, 이 서열이 (잘려나가고 남은) eIF4G에 의해 위 그림과 같이 인식될 수 있게 됨. 그 결과 정상적인 circular mRNA가 형성되고 translation이 잘 일어나게 됨.
이러한 system은 그 자체로도 흥미롭지만, 사실 우리가 실험 과정에서 이 IRES라는 서열을 이용할 수 있기 때문에 더더욱 흥미로움.
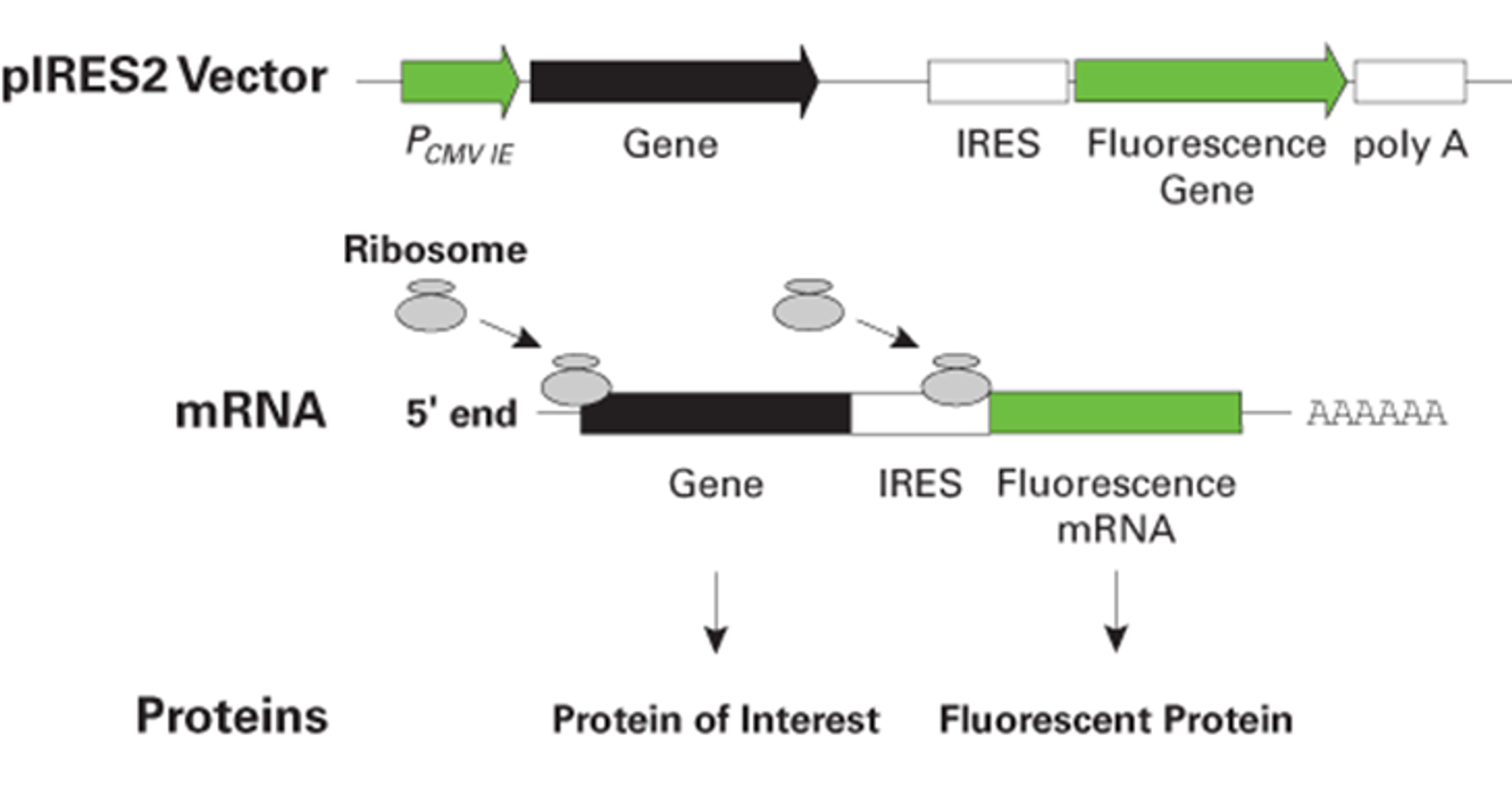
위와 같이 mRNA 중간에 IRES를 집어넣어주게 되면 진핵생물에서도 한 mRNA로부터 여러 종류의 단백질을 동시에 발현시킬 수 있음. (즉, 원래는 monocistronic한 진핵생물 system에서 polycistronic한 effect를 유발할 수 있음) 이렇게 해 줄 시 특정 gene으로부터 발현되는 단백질 산물의 expression level을 다른 gene으로부터 발현되는 단백질 산물 양으로부터 추정하는 것이 가능함. (둘이 동일한 mRNA에 있게 되면 결국 이 둘의 expression level을 같게 취급할 수 있음) 이 때문에 IRES를 사용한 polycistronic vector는 주로 antibiotic selection, fluorescence microscopy, flow cytometry를 이용한 실험기법을 이용한 expression level 측정 시 많이 활용됨.
한편 mRNA의 5' cap은 아래와 같이 많은 역할이 있다는 사실을 앞서 배웠었음.

그런데 이 때 원래는 5' cap을 인식하는 단백질로 cap-binding complex(CBC)가 붙어있는데, 이후 번역이 이루어질 때는 eIF4E와 같은 단백질이 5' cap에 붙어있음. 이 때의 exchange는 생물학적으로 꽤나 흥미로운 의미가 있다는 것이 알려져 있음.

위 그림을 보면 처음에는 CBC가 5' cap에 붙어있으면서 ribosome complex를 불러모은 뒤 시범삼아 translation을 한번 시켜봄. 이 때 단백질이 잘 만들어지고, 끝까지 만들어진다는 confirm이 이루어지고 나면 CBC는 떨어지고 eIF4E가 5' cap에 붙으면서 앞서 살펴봤었던 complex를 형성해 결과적으로 반복적인 translation이 일어나게 됨.
다음 포스트에서는 나머지 translation initiation factor들에 대해 알아보도록 하자.