
지난 포스트에서는 위와 같은 다양한 보편전사인자들이 어떤 순서로 와서 preinitiation complex를 형성하는지를 살펴봤었음.
앞서의 모식도에서 파란 구슬로 묘사된 TFIID에 대해 조금 더 자세히 알아보자. 사실 이 녀석도 다양한 subunit들로 구성됨.
여러 subunit들 중 하나가 TATA-box binding protein(TBP)임. TBP는 진화적으로 매우 보존되어 있는데, 예를 들어 yeast와 human의 TBP amino acid sequence를 비교해 볼 시 거의 80% 정도가 일치함.
한편 TBP는 신기하게도 TATA box의 minor groove에 결합함. 그렇다면 어떻게 TBP가 minor groove에 결합한다는 사실을 알아냈을까.

이에 대한 확인을 위해 위 그림 (a)에 나타난 것과 같은 substitution을 이용함. 보면 T를 C로, A를 I로 바꾸어 줬는데, 이 때 아래쪽이 minor groove를 이루는 쪽이고 위쪽이 major groove를 이루는 쪽임. 그런데 T를 C로, A를 I로 바꿔주게 되면 major groove 쪽 구조는 완전히 다르게 바뀌는 반면 minor groove는 그대로임.
TATA box 부분의 서열에서 T→C로, A→I로 substitution시켜준 후 gel mobility shift assay를 수행한 결과가 (b)에 나타나 있음. 보면 CICI일 때와 TATA일 때 TBP에 의한 binding shift 정도가 거의 동일하게 나타나는 것을 알 수 있음. 즉, TBP는 minor groove에 결합한다는 결론을 내릴 수 있음. (참고로 NS는 negative control임)
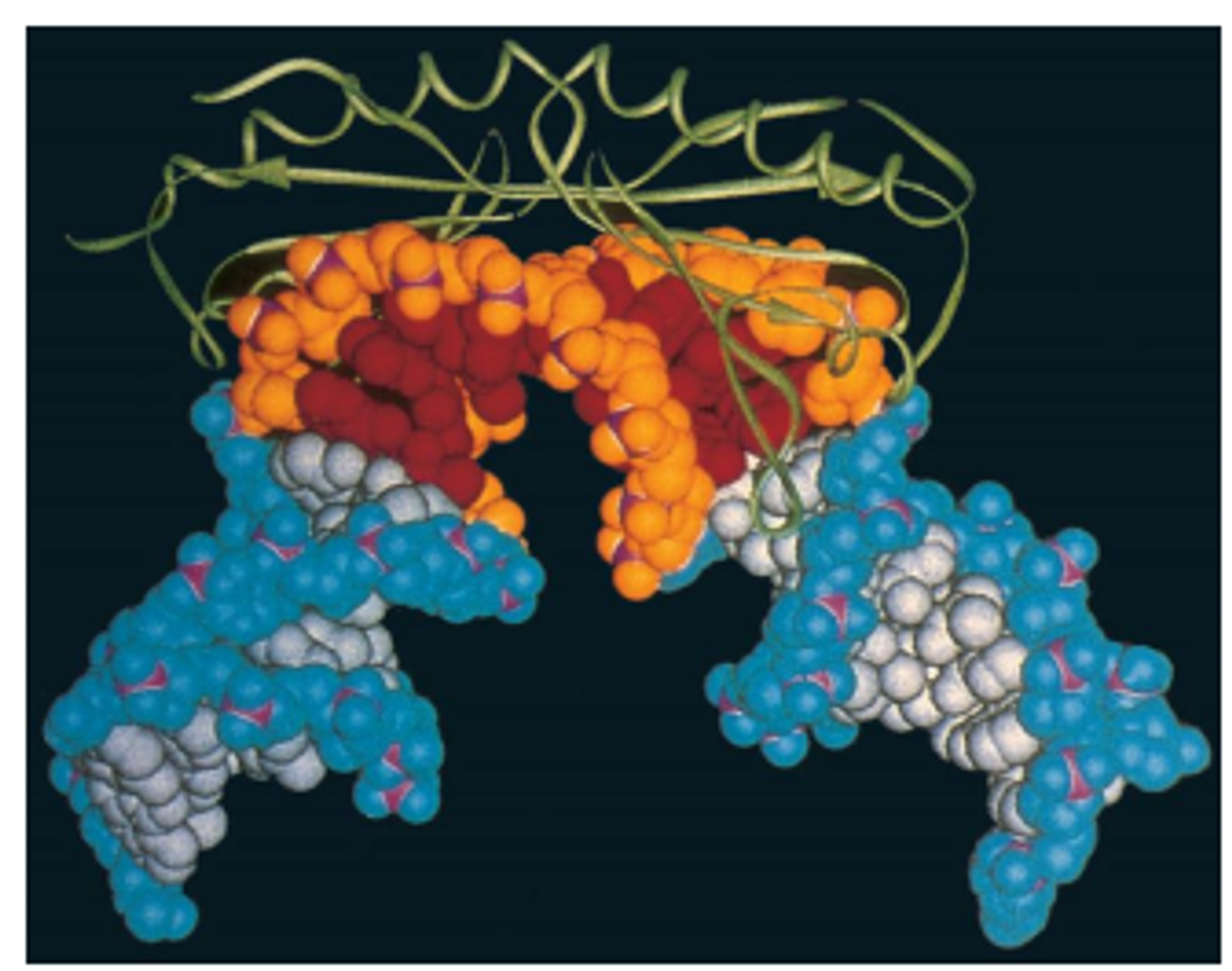
위 그림에는 실제로 TATA box(주황색)에 TBP(녹색)가 붙어있는 상태의 입체구조가 나타나 있음. 이 때 위 그림상 TATA box의 위쪽, 즉 minor groove에 TBP가 결합하고 있으며, TBP가 loop를 뻗어 TATA box와 단단히 결합하고 있음을 확인할 수 있음. 한편 위 그림에도 나타나 있듯이 TBP에 의해 TATA box 근방의 DNA가 대략 80도 정도 굽어지게 되는데, 이 bending에 의해 보다 전사가 더 잘 일어날 수 있는 형태로 변하게 됨.
참고로 이 때의 TBP 모양을 일컬어 Saddle-shape(말안장 구조)이라 지칭하기도 함.
지금까지 TBP가 마치 class II promoter에 작용해 RNA polymerase II에 의해 일어나는 전사만 돕는 것처럼 묘사되었지만, 사실 TBP는 이외의 RNA polymerase들에 의해 일어나는 전사와도 관련되어 있음. (그렇기에 TBP는 꽤나 versatile함)
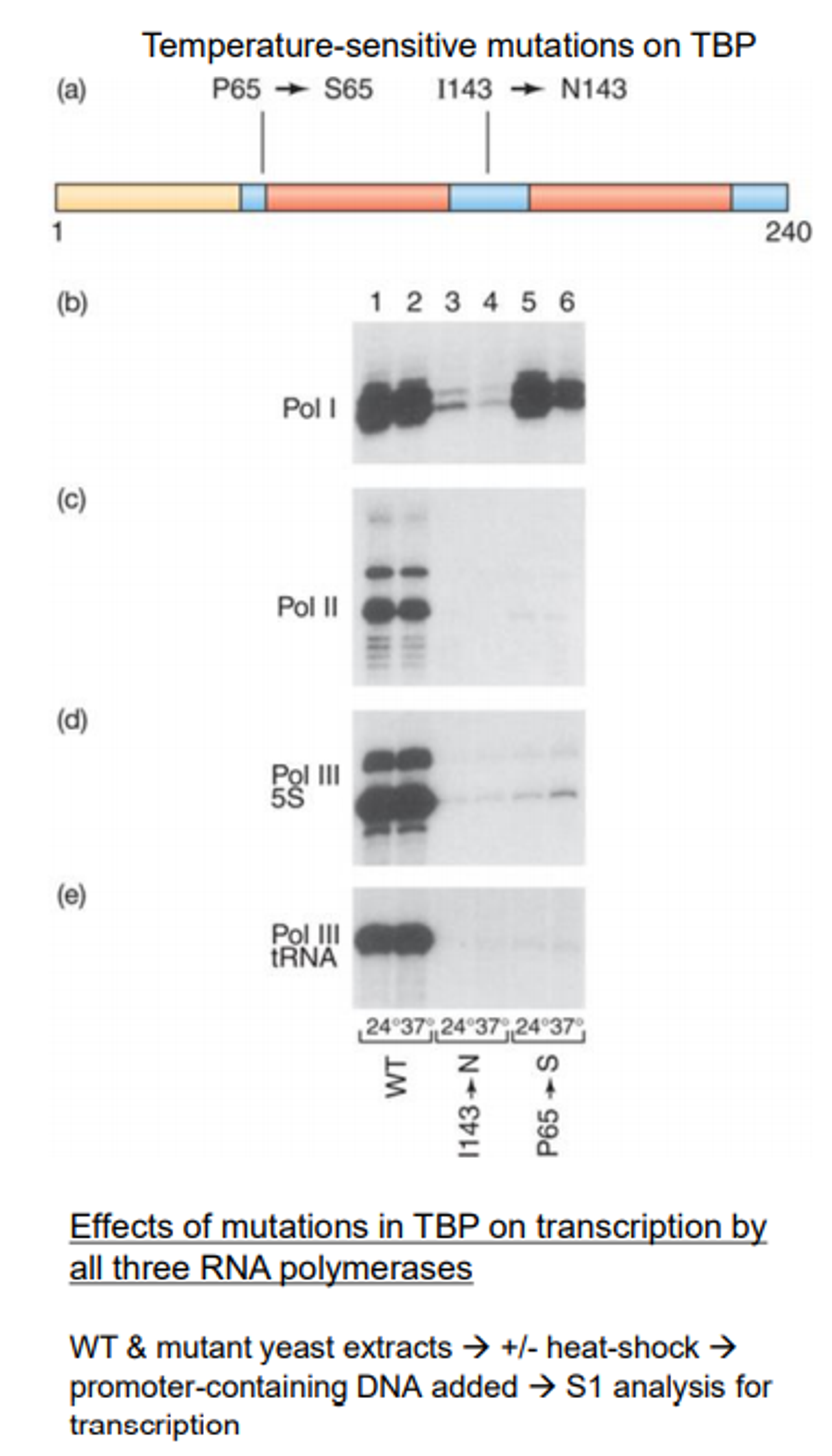
이와 관련된 실험 결과가 위와 같음. 이 실험은 위 그림의 (a)에 나타난 것처럼 TBP 중 파란색 영역의 일부를 다른 것으로(P65→S65, I143→N143) 치환해준 다음 RNA polymerase I, II, III 각각에 의한 전사가 어떤 식으로 달라지는지를 살펴봄. 그랬더니 위 그림에 나타난 것과 같이 RNA polymerase I, II, III에 의해 일어나는 전사가 TBP mutation에 의해 다 억제됨을 확인할 수 있음.
참고로 이 때 TBP에 발생시켜준 mutation은 temperature-sensitive mutation임. 이 때문에 24도에서의 결과보다 37도에서의 결과가 더 dramatic하게 나타남. (특히 RNA polymerase I에서의 3, 4 lane에서 이러한 경향성이 더 극명하게 나타남)
한편 진핵생물이 아닌 고세균(Archaea)에서도 TBP homologous가 나타나며, 고세균의 전사에 있어 중요한 역할을 수행함. 이는 고세균과 진핵생물 사이의 진화적 유연성을 보여주는 여러 증거 중 하나임.
한편 TFIID에는 TBP 뿐 아니라 13개의 TBP-associated factors(TAFs)가 존재함. 이들이 특별히 class II promoter에 결합하는 녀석들이므로 이들을 TAFII로 표현함. (다시말해 TBP와 13개의 TAFII가 모여서 온전한 TFIID를 구성하게 됨)
처음에는 13개의 TAF들을 molecular mass에 따라 명명함. 그러나 organism에 따라 TAF의 molecular mass가 달라진다는 문제점이 발생해 결국은 큰 순서대로 1~13번까지 단순히 번호로 이름붙임.
TAF는 core promoter element나 gene-specific transcription factor 들과 interaction함. 따라서 TBP와 함께 TAF가 붙으면 TBP가 붙는 TATA box 영역보다 더 확장된 DNA-protein interaction 영역이 생기게 됨. (즉, TAF는 DNA와의 interaction이 더 넓게 일어나게 도와줌)
아래 나타난 실험 결과는 TAF가 있을 때 전사 level이 어떻게 변화하는지를 나타내주고 있음.
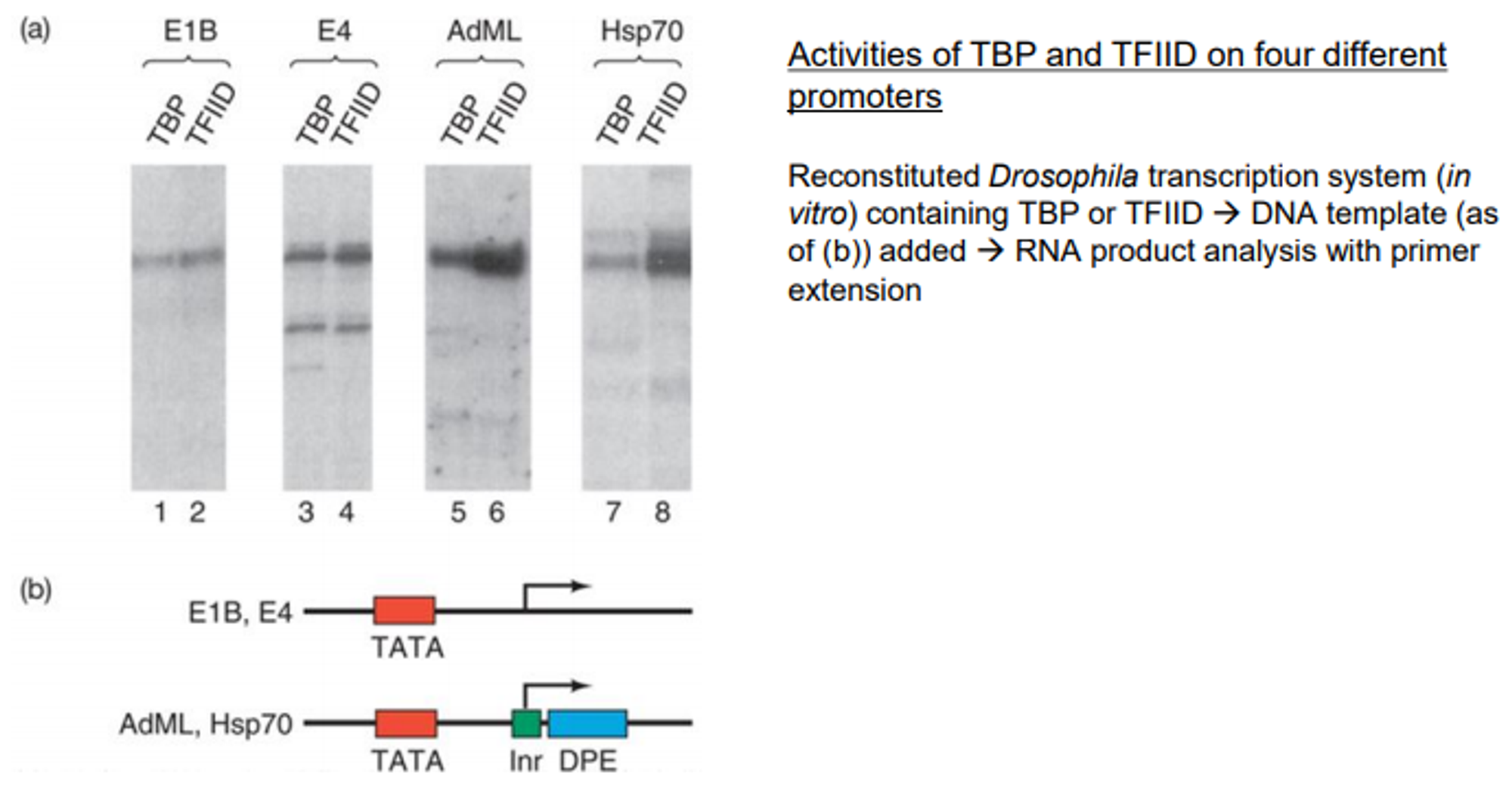
이 때 위 그림 (b)에 나타나 있는 것처럼 E1B와 E4는 TATA box만 존재하는 promoter, AdML과 Hsp70은 TATA box와 함께 Inr, DPE도 존재하는 promoter임.
(a)에 나타난 결과를 살펴보자. 우선 E1B와 E4을 promoter로 가지는 gene에서는 TBP만 있을 때와 TBP+TAF일 때의 전사 level이 거의 비슷함. 반면 AdML과 Hsp70을 promoter로 가지는 gene에서는 TBP만 있을 때에 비해 TBP+TAF일 때의 전사 level이 훨씬 더 높아짐.
이 결과를 통해 TBP는 TATA box에 binding해 전사를 촉진하고, TAF는 TATA box 이외에 Inr, DPE와 같은 다른 promoter element들과 interaction하여 전사를 촉진시킨다는 것을 알 수 있음.
다음 포스트에서 TFIID의 구조와 기능에 대해 이어서 알아보자.
'전공자를 위한 생물학 > 분자생물학' 카테고리의 다른 글
| [분자생물학] 11.3 : TFIIB의 구조와 기능 (0) | 2023.12.17 |
|---|---|
| [분자생물학] 11.2 : TFIID의 구조와 기능 - 2 (0) | 2023.12.17 |
| [분자생물학] 11.1 : 진핵생물의 보편전사인자(general transcription factor) (0) | 2023.12.16 |
| [분자생물학] 10.4 : 진핵생물의 enhancer, silencer (0) | 2023.12.16 |
| [분자생물학] 10.3 : 진핵생물의 promoter - 2 (0) | 2023.12.16 |