이번 포스트부터는 lac operon(젖당 오페론)에 대해 알아보자.
lac operon은 operon 중 처음으로 발견된 종류에 해당함. 일반적으로 operon이라는 개념은 원핵생물에게서만 존재하는 개념으로, 하나의 promoter에 의해 여러 gene들이 하나의 mRNA로 전사된 후, 각각의 gene으로부터 유래한 mRNA의 서열들이 서로 독립적으로 ribosome과 결합해 발현하게 되는 방식임. (이러한 원핵생물의 특성을 polycistronic하다고 표현함)
E.coli가 가지고 있는 lac operon에는 3가지 gene이 포함되어 있는데, lacY, lacZ, lacA가 그것임. lacY는 galactoside permease를 encoding하고 있으며 lactose를 cell 내부로 transport하는 역할을 함. 다음으로 lacZ의 경우 beta-galactosidase를 encoding하고 있으며 lactose를 galactose와 glucose로 분해시켜주는 역할을 함. 마지막으로 lacA의 경우 galactoside transacetylase를 encoding하고 있으며 이 녀석의 역할은 아직 불분명함.
아무튼 이 lac operon은 glucose가 고갈된 후 lactose만 남았을 때 lactose를 에너지원으로 가공하기 위해 on되게 되며, 그 결과 lacY, lacZ, lacA가 발현되어 lactose의 분해를 도와주게 됨. (lac operon에 의해 발현되는 단백질들이 결과적으로 lactose의 '분해'를 도와주므로 이들 효소들을 catabolic enzyme이라 부르기도 함)

보면 glucose가 사용된 이후 어느정도의 정체기를 거쳐 lactose가 사용되고 있다는 것이 위 그림 왼쪽 graph에 나타나 있음. 이 정체기 동안 지금부터 배울 lac operon의 발현이 일어나게 됨. 참고로 위 그림 오른쪽에는 lactose가 beta-galactosidase에 의해 galactose와 glucose로 분해되는 기작이 나타나 있음. (결국 이 기작이 lac operon의 켜에 의해 유도되게 되는 것)
lac operon은 매우 정교하게 control되고 있음.

일반적으로 lactose가 없을 때는 lac operon이 발현되지 않는데, 이 때 repression의 과정이 (a)에 나타나 있음. 일단 위 그림상에서 lacZ, lacY, lacA로 표시된 부분이 실제로 발현되는 structural gene에 해당하며, 그 앞쪽에 operator, promoter가 존재함. (operator보다 promoter가 더 upstream에 있다는 것을 유의할것)
한편 promoter보다 더 upstream에는 lacI라고 하는 gene이 operon과는 별개로 계속 발현하고 있음. 이 때 이 녀석의 발현에 의해 만들어지는 protein은 repressor임.이 repressor monomer 4개가 모여 그림상에 나타난 것처럼 tetramer를 형성하게 되면 이 repressor tetramer가 operator 부위에 붙게 되고 그 결과 RNA polymerase에 의한 전사가 일어나지 않게 됨. (즉, lacZ, lacY, lacA가 발현되지 않음)
한편 (b)에서 나타난 상황과 같이 lactose가 존재하고 있을 경우 lactose의 양 증가에 따라 inducer의 농도도 같이 증가하게 됨. 이 때 inducer는 repressor tetramer에 allosteric하게 결합하게 되며, 그 결과 repressor의 conformation change에 의해 repressor가 operator에 결합하지 못하게 됨. 그러면 결국 RNA polymerase에 의한 전사가 잘 일어나 결과적으로 lacZ, lacY, lacA가 발현됨.
그렇다면 lactose 증가에 따라 양이 같이 증가하는 inducer의 정체는 도대체 무엇일까.
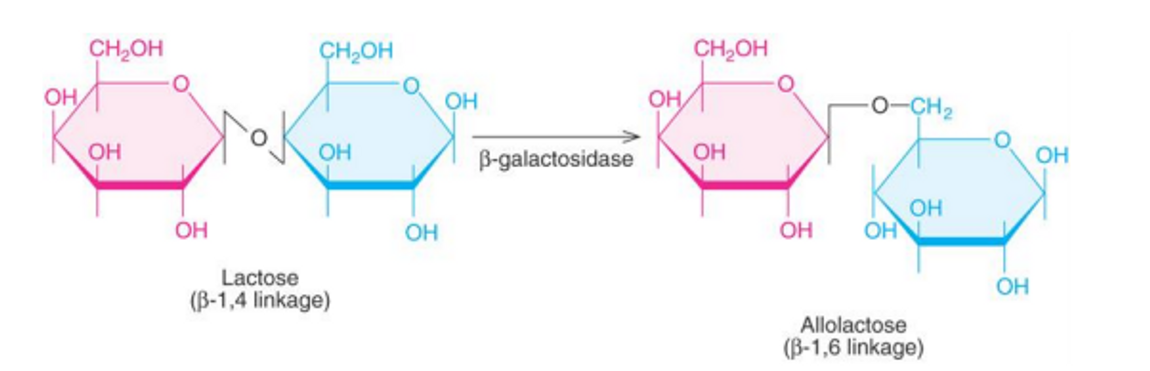
위 그림과 같이 lactose는 beta-galactosidase에 의해 allolactose로 변환될 수 있는데, 이 때 이 allolactose가 바로 inducer로 기능함.
(참고로 이 때 lactose의 존재에 의해 \beta-galactosidase를 encoding하는 gene인 lacZ의 발현이 촉진되어야 하는데, 어떻게 inducer를 생산하는 위 과정에서 beta-galactosidase가 사용되는 것이 가능한가에 대해 의문이 생길 수도 있음. 실제로는 lac operon이 꺼져 있을 때에도 beta-galactosidase는 소량 발현되고 있고, 소량 발현된 beta-galactosidase만으로도 lactose를 allolactose로 바꿔주기에는 충분하므로 일단 lactose가 존재하기만 하면 inducer인 allolactose의 전환에는 전혀 무리가 없음. 이후 allolactose에 의해 repressor의 활성이 저해되면 그 결과 lac operon이 활성화되어 beta-galactosidase의 합성이 대량으로 일어나게 됨)
그렇다면 operon은 어떻게 발견된 것일까. 1940~1950년대에 Jacob와 Monod는 E. coli에서의 lactose metabolism에 대해 연구하고 있었는데, 이 과정에서 operon에 대한 증거를 찾게 됨.

위 표는 그때 당시의 발견 결과를 보기 쉽게 정리한 것임. 이 때 Z+는 lacZ가 정상기능을 하는 것을, Y+는 lacY가 정상기능을 하는 것을 의미하며, 이 때 accumulation을 관찰한 galactoside는 lactose의 일종임. (따라서 Y^+인 조건에서 galactoside는 세포내부로 수송되어 들어올 것임) 이 때 Z+Y+인 조건에서, inducer를 넣어주지 않았을 때는 glalactoside가 세포 내에 축적되지 않았고 inducer를 넣어줬을 때는 축적됨. 이를 통해 (아직 구체적인 정체는 몰랐지만) inducer에 의해 galactoside를 세포 내로 들어오게끔 하는 유전적 switch가 켜질 수 있음을 알게 됨. (즉, 이에 관여하는 정체모를 gene이 있으며 이 gene의 발현을 inducer가 조절할 것이라 추측함) 한편 Z+Y-의 genotype을 가지는 개체의 경우 inducer를 넣어주던 넣어주지 않던 galactoside가 세포 내에 축적되지 않았음. 이를 통해 lacY가 galactoside를 세포 내로 들어오게 해주는 유전자임을 확인함.
다음 포스트에서는 이어서 operon과 관련된 다양한 mutant들에 대해 알아보자.
'전공자를 위한 생물학 > 분자생물학' 카테고리의 다른 글
| [분자생물학] 7.1 : lac operon(젖당 오페론) - 3 (0) | 2023.08.31 |
|---|---|
| [분자생물학] 7.1 : lac operon(젖당 오페론) - 2 (1) | 2023.08.31 |
| [분자생물학] 6.4 : 원핵생물의 transcription termination(전사 종결) (0) | 2023.08.30 |
| [분자생물학] 6.3 : 원핵생물의 transcription elongation(전사 신장) (0) | 2023.08.30 |
| [분자생물학] 6.2 : 원핵생물의 transcription initiation(전사 개시) - 5 (0) | 2023.08.30 |