이번 포스트에서는 텔로미어와 텔로머레이스에 의한 작용에 대해 자세히 알아보자.
telomere는 TTAGGG reapeat으로 이루어져 있음. 이 repeat은 사람, 쥐에게서 동일하게 나타나며, 사람의 경우에는 대략 10kb 정도의 길이만큼 repeat이 이루어져 있음. (쥐의 경우 50kb정도)

한편 repeat 뒷부분에는 G-rich strand라는 부분이 대략 50-300nt 길이로 존재하고 있는데(ssDNA) 이 서열의 경우에는 telomerase에 의해서 유지됨.

telomerase의 작동 과정에 대한 모식도가 위 그림에 나타나 있으므로 참고할 것.
telomere는 chromosome의 end-protection problem을 해결하는데 핵심적인 도움을 줌.
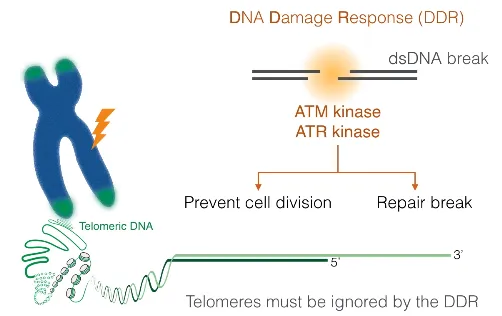
위 그림에 나타나 있는 것과 같이 원래 dsDNA가 break되면 ATM kinase, ATR kinase 등에 의해서 DNA damage response(DDR)가 일어남. 그런데 telomere도 사실 break가 일어난 dsDNA와 동일한 모양새임. 그러나 DDR을 유발시키지는 않음. 어떻게 이게 가능할까.
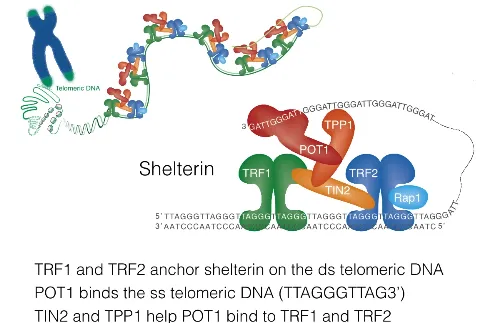
telomere에는 위와 같이 단백질 complex가 결합함. dsDNA를 인식하여 결합하는 TRF1, TRF2 뿐만 아니라 ssDNA를 인식하여 결합하는 POT1과 같은 SSBP protein 등이 모두 모여서 총 6개의 protein으로 이루어진 Shelterin complex를 형성함.

위 그림에는 dsDNA를 인식해 결합하는 TRF1, TRF2에 대한 모식도가 나타나 있음. 이들은 dimer를 형성하는 한편, Myb이라고 하는 triple-helix 구조를 가져서 이 motif로 telomere 서열을 인지함.
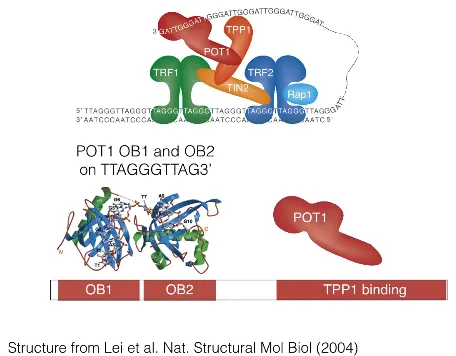
한편 위 그림에는 POT1이 나타나 있음. 이 녀석은 N-terminal 부분에 OB1, OB2와 같은 domain을 가지고 있으며, 이 부위에 의해서 ssDNA를 인지할 수 있음. (3' 말단을 인지할수도, 중간부분을 인지할수도 있음)

위 그림의 우측에 ATM pathway가 나타나 있음. 보면 DNA end 부분을 인지한 MRN이라는 단백질이 ATM kinase를 recruit하고, ATM kinase에 의해서 DNA damage foci를 marking해주는 MDC1, 53BP1과 같은 단백질들이 recruit됨. (이 단백질들이 일종의 DNA damage foci marker) 한편 ATM kinase는 인산화 기작에도 관여해 결과적으로 p53의 활성은 높이고 Cdc25의 활성은 낮춰서 cell이 G1에서 S기로 넘어가지 못하게 block시켜버림. 그런데 이 전반적인 ATM pathway를 Shelterin이 inhibition시켜버림.
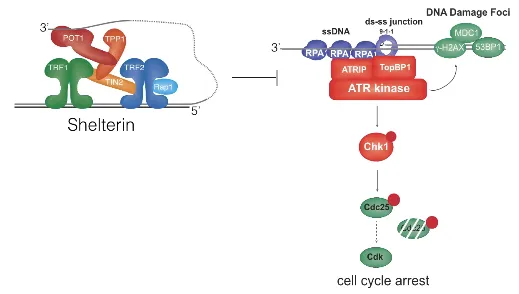
한편 위 그림의 우측에는 ATR pathway가 나타나 있음. 이 경우 ssDNA를 인지하고 결합하는 RPA(서열 상관없이 ssDNA이기만 하면 결합함)에 의해서 ATR kinase가 recruit되고 이후 여러 DNA damage foci marker protein들이 차례대로 recruit되는 한편 ATR kinase에 의해서 인산화 relay가 일어난 결과 cell cycle의 arrest가 일어남. 이러한 과정을 Shelterin이 inhibition시켜버림.
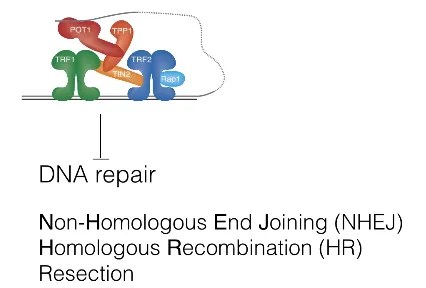
한편 Shelterin은 위 그림에 나타나 있는 것과 같이 DNA repair 과정을 block하기도 함.
그렇다면 도대체 어떻게 단지 6개의 subunit으로 이루어진 Shelterin이라는 단백질이 이렇듯 다양한 조절을 할 수 있는 것일까. 이에 대해 연구하기 위해 mouse embryo fibroblast(MEF)를 이용한 실험을 수행할 수 있음. 물론 아예 Shelterin을 deletion시켜버리면 이는 lethal하므로, 아래와 같은 conditional gene deletion method를 사용함.
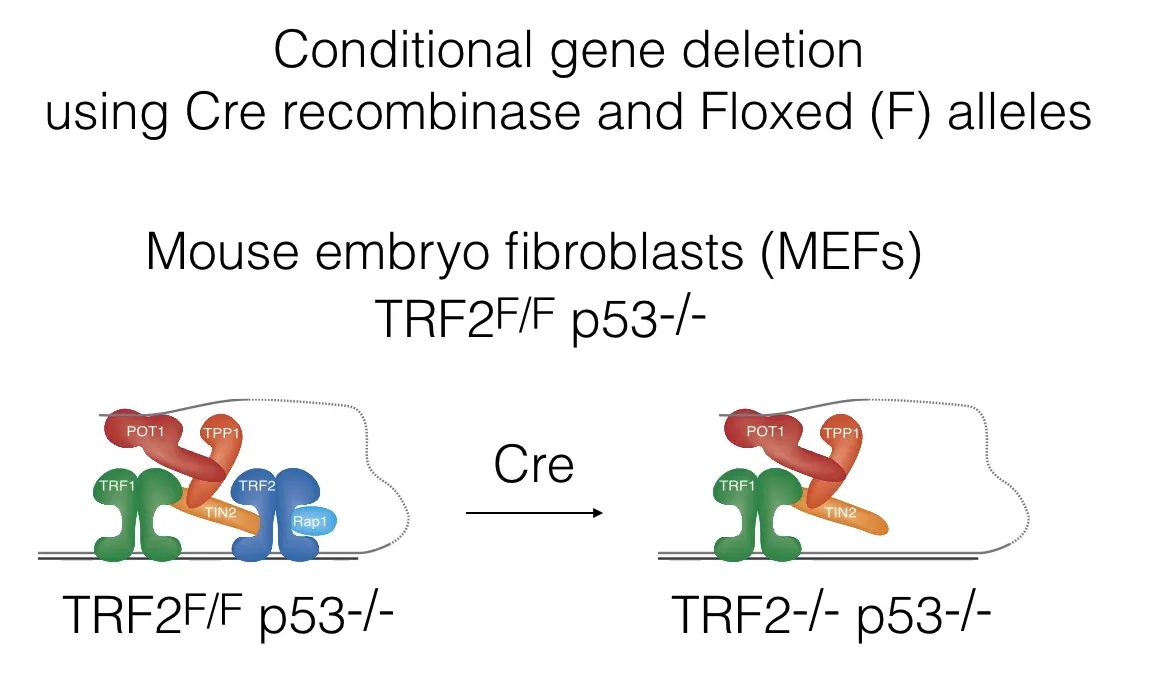
이 때 사용한 것은 Cre recombinase와 Cre recombinase에 의해 잘릴 수 있는 서열임. 구체적으로는 Shelterin encoding gene 중 TRF2 부분 사이에 인위적으로 Cre recombinase에 의해 잘릴 수 있는 Cre 서열을 넣어준 후(Floxed allele) Cre recombinase를 가해줌. 그 결과 위 그림 아래와 같이 Shelterin 중에서 TRF2만 없앨 수 있었음. (참고로 이 때 p53도 같이 빼준 이유는 p53이 있을 시 apoptosis가 유발되어버리지만 없을 시 TRF2를 없앴을 때의 telomere 표현형을 지속적으로 관찰할 수 있기 때문)
일단 위와 같이 TRF2를 없애줄 시 TFR1에 의해서도 TIN2, POT1, TPP1과 같은 녀석들이 recruit되는데는 전혀 문제가 없다는 것을 알 수 있음. 그 밖에 TRF2에 의해서 recruit되는 Rap1의 경우에는 당연히 TRF2가 없을 시 recruit되지 않음. 다만 추가적인 연구를 통해서 Rap1은 Shelterin activity에 있어서 크게 영향을 미치지 않는다는 것이 확인되었으므로 결과적으로 위 그림에서 만든 mutant의 phenotype을 관찰하면 TRF2의 역할을 짐작할 수 있음.
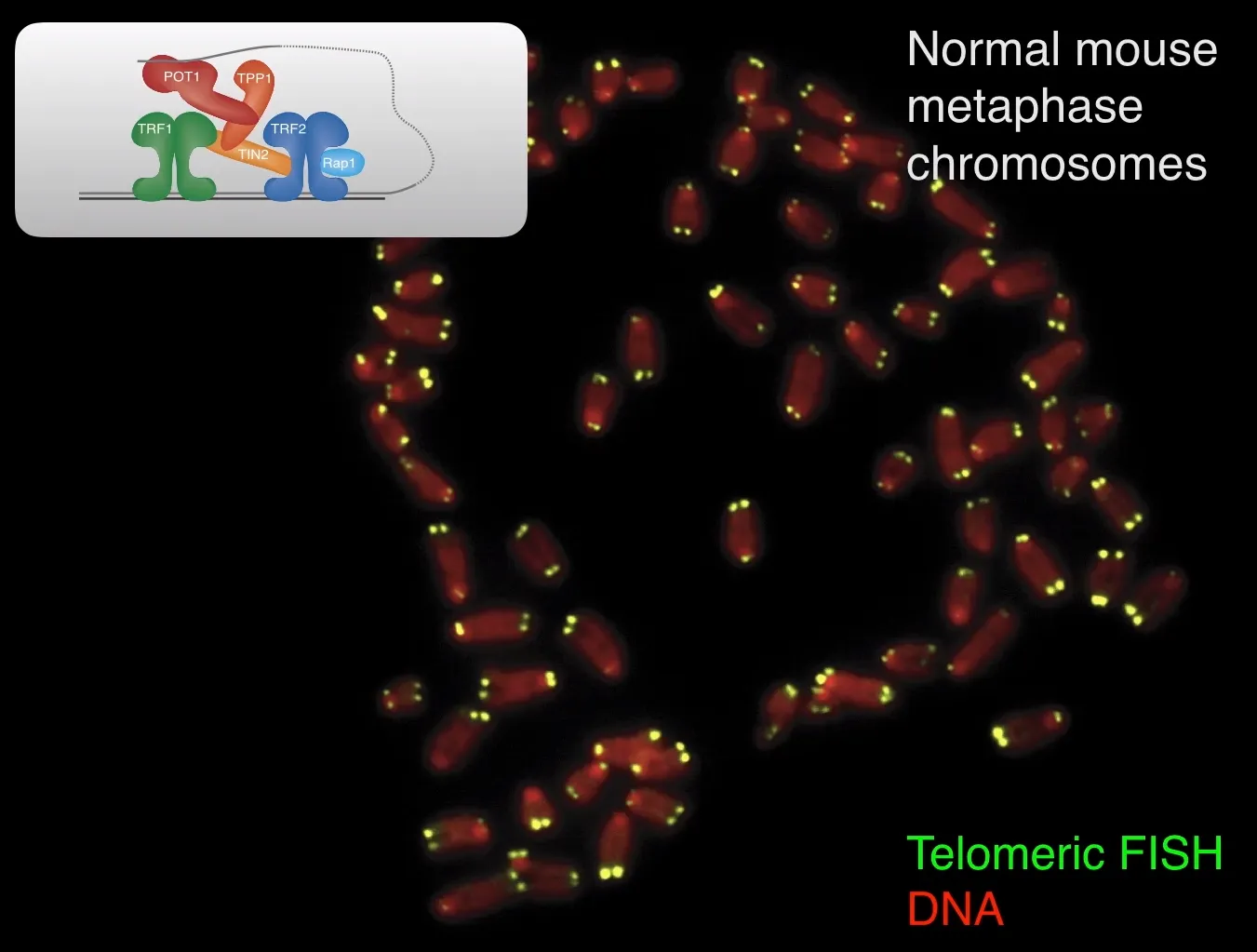
우선 위 그림은 정상적인 mouse cell에서의 metaphase에서 telomeric FISH를 이용해 녹색으로 telomere를 표지한 것임.
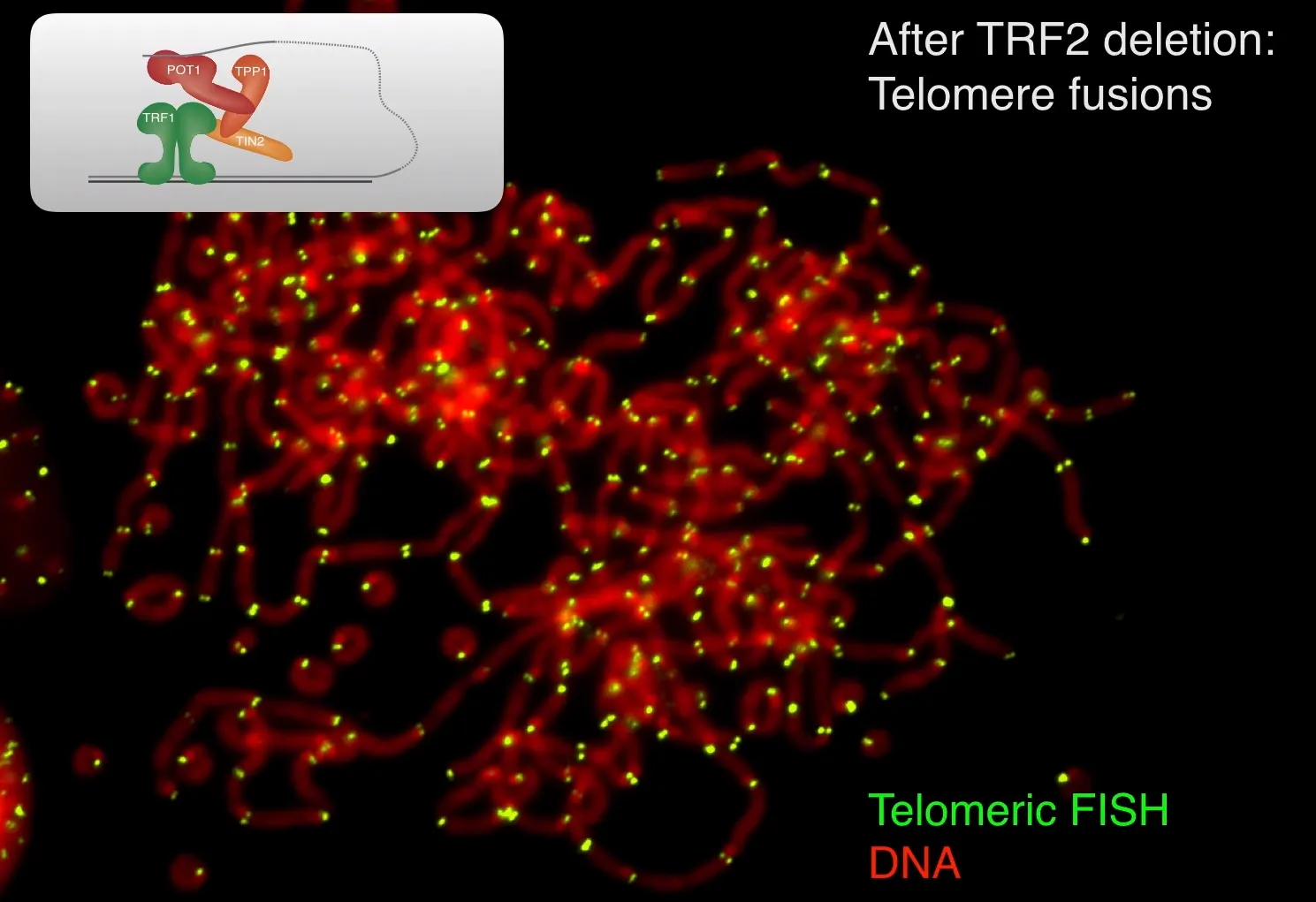
한편 위 그림은 TRF2 deletion을 시킨 mouse cell에서의 metaphase에서의 telomere 표지 결과임. 보면 DNA 중간에 telomere가 존재하고 있으며, 서로서로 얽혀있는 등 굉장히 이상한 구조를 하고 있음을 확인 가능함. 이는 telomere의 끝부분이 repair로부터 보호되지 못해서 partial한 DNA repair가 나타난 결과임. 특히 non-homologous end joining(NHEJ)에 의해서 telomere간의 fusion이 일어남.
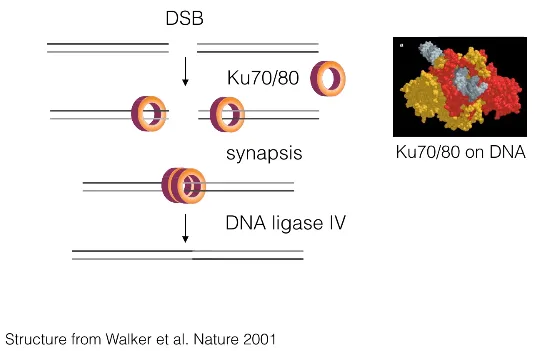
NHEJ의 기본적인 작동과정은 위와 같음. 보면 ring shape를 가지고 있는 Ku70/80 heterodimer가 끊어진 DNA에 딱 맞게 들어감. (끊어지지 않은 DNA에는 이 녀석이 들어갈 수 없음) 이후 Ku70/80끼리 synapsis를 통해 합쳐지고 이어서 DNA ligase IV가 recruit되면서 서로 끊어져있던 두 DNA들이 연결되게 됨.
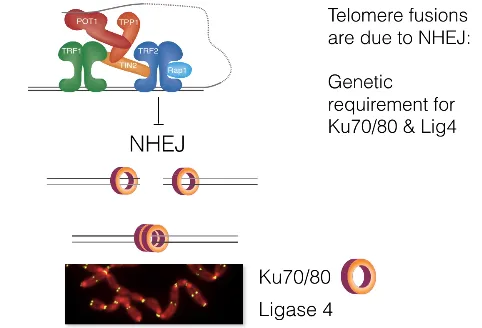
이런 NHEJ의 과정을 원래는 TRF2가 block하고 있음. 이에 대해 알게 된 것은 TRF2 deletion mutant에게 Ku70/80 혹은 ligase 4를 제거해봤을 때 앞서와 같은 telomere fusion이 생기지 않는다는 것을 확인하고 난 뒤임. 아무튼 결론적으로 Shelterin 중에서도 TRF2가 NHEJ를 block하는 역할을 담당한다는 것을 알 수 있었음.
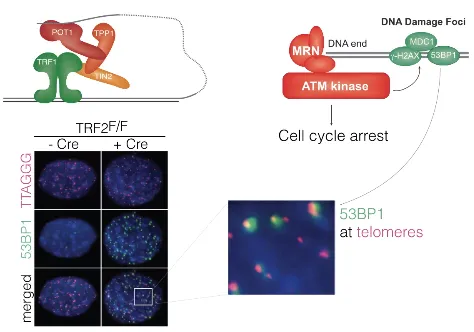
한편 위 그림에 나와있는 실험을 통해서 TRF2가 ATM pathway도 block하는 역할을 한다는 것을 알 수 있음.
보면 Cre recombinase가 있어서 TRF2가 제거된 경우에서만 telomere와 53BP1(ATM kinase에 의해 recruit되는 DNA damage foci의 marker protein 중 하나)가 동일한 위치에 염색됨. 이를 통해 원래는 TRF2가 ATM kinase에 의한 DNA damage foci marking을 block하고 있었다는 것을 알 수 있음.
다음 포스트에서는 이번 포스트에 이어서 telomere의 형성 과정 및 조절 과정에 대해 알아보도록 하자.
'전공자를 위한 생물학 > 분자생물학' 카테고리의 다른 글
| [분자생물학] 21.1 : DNA damage와 수선 기작 (0) | 2024.10.29 |
|---|---|
| [분자생물학] 20.3 : 텔로미어와 텔로머레이스 - 2 (1) | 2024.10.29 |
| [분자생물학] 20.2 : DNA 복제 종결 (0) | 2024.10.27 |
| [분자생물학] 20.1 : DNA 복제 개시 - 2 (0) | 2024.10.27 |
| [분자생물학] 20.1 : DNA 복제 개시 - 1 (0) | 2024.10.27 |