이번 포스트에서는 단백질 항상성(proteostasis)에 대해 알아보자.
proteostasis는 protein homeostasis를 의미함.
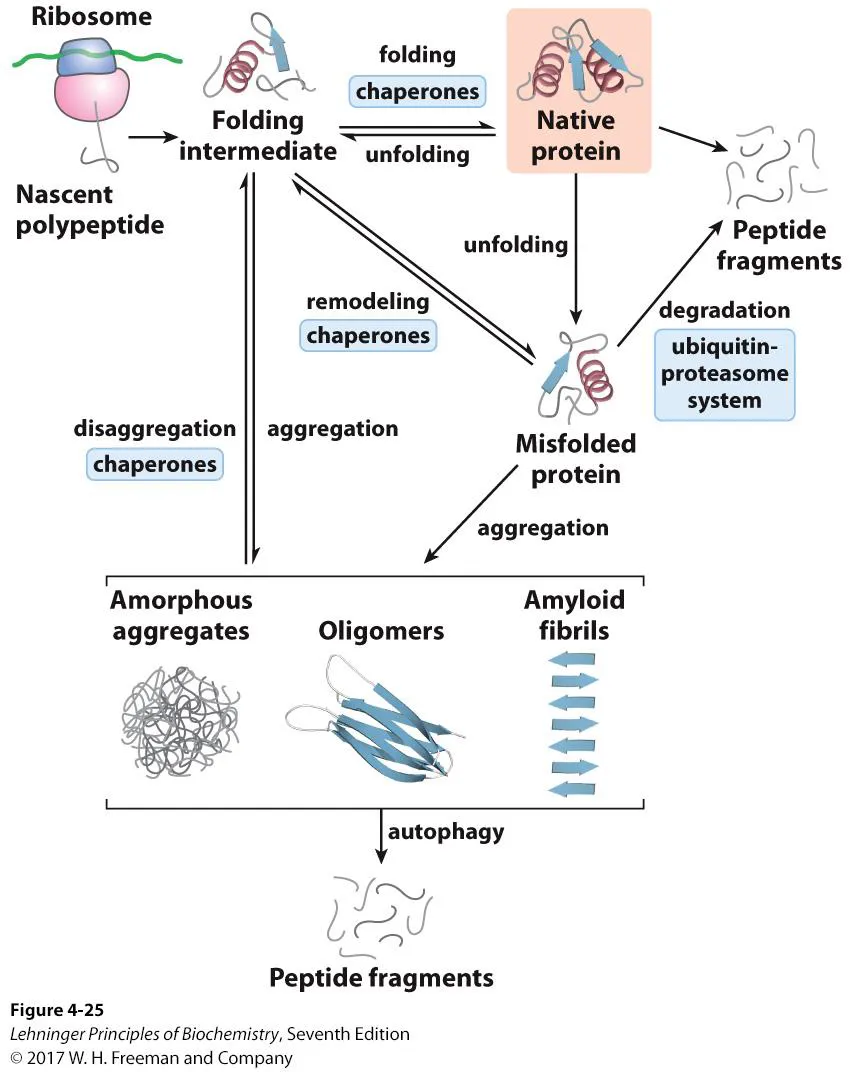
위와 같이 ribosome에서 처음 만들어진 protein은 별다른 구조를 형성하고 있지 않음. 그러다가 이 녀석이 folding되기 시작하는데, 그 과정에서 일부 단백질들은 저절로 native protein으로 folding되기도 하지만 대부분의 경우 chaperone의 도움을 받아 folding되게 됨. 한편 native protein도 때에 따라 unfolding될 수 있는데, 그렇게 해서 만들어지는 것이 바로 misfolded protein임. (native protein 구조의 일부가 풀려서 나타나므로 이상한 모양일 것임) 이 때 이들의 misfold 정도가 심하지 않을 경우 chaperone에 의해 다시 remodeling되어서 native protein으로 refolding되기도 하고, misfold 정도가 심하다면 ubiquitin-proteasome system(UPS)을 통해서 peptide fragment로 분리되기도 함.
그러나 일부 misfolded protein들은 자기들끼리 aggregation되어서 amorphous aggregate(특정한 구조가 없이 그냥 뭉처진 형태), oligomer, amyloid fibril 등을 형성하기도 함. 이런 녀석들은 온갖 질병을 유발할 수 있음.
(물론 이런 녀석들을 어느정도 제거해줄 수 있는 autophagy라는 기작도 존재하기는 함)
protein의 기능은 protein의 3D structure에 따라 결정된다고 보아도 과언이 아님. 실제로 native fold 형태가 가장 functional하며 여기서 만약 unfolding이 일어난다면 그 결과 denaturation되어서 activity를 점차 잃어갈 수 있음.
denaturation을 시킬 수 있는 가장 대표적인 조건들이 heat, cold, pH extreme, organic solvent(이들 중에서 특히 물과 안 친한 녀석들을 사용할 경우 protein 중 hydrophobic한 녀석들이 되려 바깥쪽으로 나온 결과 denaturation이 됨), chaotropic agent(urea, guanidinium hydrochloride) 등임.

실제로는 위와 같은 titration을 통해 단백질의 unfolded 된 정도(다시말해 단백질의 stable한 정도)를 측정할 수 있음. 우선 왼쪽 graph를 보면 온도가 높아짐에 따라서 protein들이 어떻게 unfolded되는지의 경향을 graph로 알 수 있음. (이 때 y축으로 0에 해당하는 시작부위가 바로 native fold임) 이런 측정에는 주로 CD가 사용됨.
특히 이 graph의 Tm값(melting temperature)을 비교해보면 ribonuclease A에 비해 Apomyoglobin이 더 고온에서도 stable하다는 것을 알 수 있음. (참고로 Apomyoglobin은 heme group을 안 가지고 있는 myoglobin임)
한편 위 그림 오른쪽 graph를 보면 chaotropic agent를 처리해준 경우 점차적으로 ribonuclease A의 denaturation이 유발된다는 것을 알 수 있음.
참고로 ribonuclease A는 소 등의 동물로부터 쉽게 얻을 수 있는 protein이어서 초반에 연구에 많이 사용되었었음.
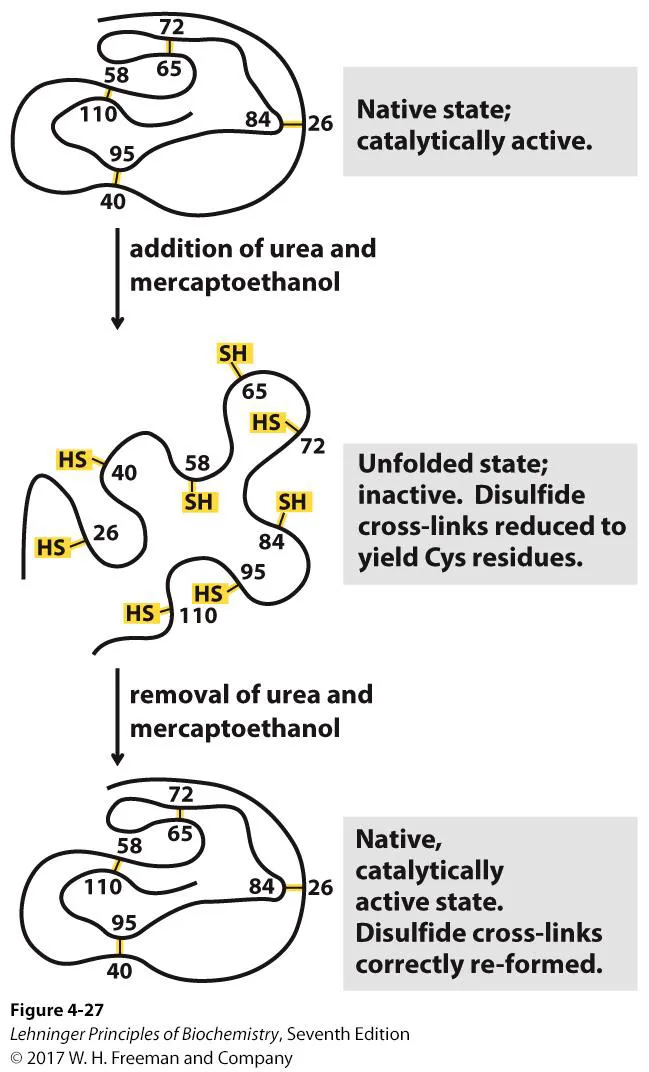
위 그림에 나타나 있는 것은 Chris Anfinsen이 ribonuclease를 이용해 수행한 refolding experiment임.
일단 ribonuclease는 위 그림 맨 위에 나타나 있는 것처럼 4개의 disulfide bond로 연결되어 있음. 따라서 이 녀석은 그냥은 매우 안정하므로 잘 denaturation되지 않고, reducing agent인 2-mercaptoethanol(β-mercaptoethanol)을 사용해서 disulfide bond를 끊어줘야 비로소 denaturation을 시킬 수 있음. (reducing agent들은 자기들끼리 disulfide bond를 형성하면서 실제 protein 내부의 disulfide bond는 끊어줌) reducing agent들을 처리해준 다음에는 열, pH, urea와 같은 자극을 줘서 protein을 denaturation시켜주게 됨. 그 다음에 urea 등의 denaturation source를 제거해주고 reducing agent도 제거해줌. 그러면 흥미롭게도 ribonuclease의 경우 chaperone 없이도 자기 스스로 다시 renaturation되면서 완벽히 native structure를 찾아감. (이 때 매우 흥미로운 것은, 수많은 S-S 결합의 가능성이 있지만 그 중 딱 native structure에 맞는 S-S pair만이 만나게 된다는 것임)
Anfingen은 이 결과를 바탕으로 protein sequence 자체가 native conformation을 결정할 수 있지 않을까 하는 생각을 하게 됨. (이 생각은 일반적으로는 틀렸지만, ribonuclease에 한해서는 맞을 수 있음)

현재는 protein이 위와 같이 folding된다고 생각함. 보면 일부 region들이 모여 먼저 특정 구조를 형성한 다음, 이미 형성된 구조들끼리 다시 interaction해서 최종적인 native structure가 만들어지게 됨.
한편 생체 내에서 protein은 거의 μs정도, 길어봐야 s 단위의 시간 안에 접힘. 그러나 생각해보면 단백질이 가질 수 있는 φ, ψ angle을 고려해봤을 때 protein 전체를 구성하는 amino acid각각이 가질 수 있는 conformation이 너무나 많아서 빠른 시간동안 접히는게 얼핏보면 불가능한듯 보임. 이러한 딜레마를 Levinthal's paradox라 함.

현재는 native structure가 만들어지는 과정을 위와 같은 Free-energy funnel model로 이해함.
우선 위 그림 a를 보자. 이 경우 위쪽의 넓은 부분이 불안정하며 dynamic한데다가 경우의 수도 많은 아직 접히지 않은 구조이며, 맨 아래에 N으로 표시된 부분, 즉 native fold가 가장 energy적으로 안정하기에 마치 unstructured protein이 깔때기 빨려들어가듯 쑥 N으로 변하게 됨.
다음으로 b를 살펴보자. 사실 이 경우가 대부분의 protein들에 적용되는 story임. 보면 이 경우에도 가장 에너지적으로 안정한 상태는 N이지만, 그 밖에 중간중간에 local minima(일종의 trap)가 있음. 따라서 protein이 접히다가 local minima에 도달하게 되면 trap되고, 그렇기에 이 상태에서는 외부에서 일정양 이상의 energy가 가해져야만 다시 N으로 갈 수 있음. 이 때 chaperone이 local minima에 trap된 녀석들이 N으로 제대로 접힐 수 있도록 도와줌.
다음으로 c를 살펴보자. 이 경우 특정한 구조적 접힘 과정이 없는 IDP가 해당함.
마지막으로 d를 살펴보자. 이 경우 꽤나 안정한 intermediate들이 존재하고 있음. 따라서 이 intermediate가 N이 되기 위해서는 사실상 chaperone의 도움이 반드시 필요함.
다음 포스트에서는 샤페론(Chaperone)과 샤페로닌(Chaperonin)에 대해 알아보자.
'전공자를 위한 생물학 > 생화학' 카테고리의 다른 글
| [생화학] 4.1 : protein과 ligand (0) | 2024.11.06 |
|---|---|
| [생화학] 3.6 : 단백질 항상성(proteostasis) - 2 (0) | 2024.11.05 |
| [생화학] 3.5 : 단백질의 구조 결정 방법 (0) | 2024.11.05 |
| [생화학] 3.4 : 단백질의 3차구조(tertiary structure) - IDP (0) | 2024.11.05 |
| [생화학] 3.4 : 단백질의 3차구조(tertiary structure) - globular protein (0) | 2024.11.05 |