이번 포스트부터는 집단유전학에 대해 알아보도록 하자.
본격적인 내용을 다루기 전에 우선 기본적인 용어의 definition에 대해서부터 짚고 넘어가자.
소진화는 동종 내에서의 형질 변화를 의미하고 대진화는 종 자체가 변화하는 것을 의미함. 한편 집단(population)은 같은 지리적 위치에 살았고 active하게, 혹은 potential하게 interbreed를 한(즉, 한 집단 내에서 교배를 한) 공통된 gene set을 지닌 individual들이 모여있는 group을 의미함. 그리고 유전자풀(gene pool)은 한 집단 안에 존재하는 모든 대립 유전자들을 의미함.
대부분의 집단들은 상당히 높은 수준의 heterozygosity(이형접합성)을 가지고 있음. 다시 말해 한 집단 내에서 모든 gene이 homozygous인 경우는 없다고 보아도 무방함. (쌍둥이조차도 태어난 후 spontaneous mutation에 의해 서로의 gene들이 달라짐)

인간들은 약 15,000년 전부터 야생 늑대에 존재하는 유전적인 다양성 중에서 원하는 특성만을 인위적으로 선택하였고(artificial selection), 그 결과 개의 가축화가 진행됨.
한편 genetic variation은 염기서열의 변이로부터 생긴다고 볼 수 있음. (그리고 이 중에서 특히 생식세포에서 일어나는 변이만이 자손에게 전달될 수 있음)
염기서열 변이에 의해 생기는 질병 중 대표적인 예로 cystic fibrosis가 있는데, 이 질병은 CFTR을 encoding하는 CF gene에 생긴 mutation에 의해 발생 가능함. (참고로 이 때 CF gene에 생길 수 있는 1,800개의 서로 다른 mutation 종류가 발견, 보고됨. 특히 전체 mutation의 60%의 빈도로 관찰되는 것이 exon 10의 3bp deletion임) 구체적으로는 CF gene에 대한 mutation이 recessive homo로 만나면 cystic fibrosis가 발병됨.
그렇다면 왜 이러한 병이 진화적으로 도태되어서 사라지지 않는 것일까. 이는 자발적인 mutation이 계속 발생하기 때문임. 이와 관련해 1968년 기무라가 한 설명이 바로 neutral theory(중립설)임. neutral theory란, 대부분의 mutation들은 효과가 neutral해서 진화압을 받아 도태되지 않고 남아있다는 이론임. (예를 들어 SNIPs의 경우 사람마다 다 다르게 mutation이 일어났지만 대부분 neutral하므로 사라지지 않고 남아있음) 한편 이 밖에 높은 수준의 유전적 변이는 natural selection(자연선택)으로도 설명됨. 예를 들어 단세포 생물의 경우 외부환경과 바로 맞닿아 있기 때문에 다양한 mutation들을 바탕으로 한 빠른 적응이 필수적이라 할 수 있음. 이러한 이유들에 의해 높은 수준의 genetic variation이 관찰되는 것임.
microevolution(소진화)은 집단 내에서 생식적 격리(reproduction isolation)가 발생하지 않으면서 대립유전자의 빈도가 변하는 현상을 의미함. 이와 관련된 상당히 유명한 law가 바로 Hardy-Weinberg law(하디-바인베르크 법칙)임. 이 법칙은 ideal한 상황에서 어떻게 gene pool의 특정 유전자 빈도가 유지되는지를 설명해줌. 그런데 실제 자연은 ideal하지 않으므로, 자연에서는 Hardy-Weinberg law이 성립하지 않게 되고, 그 결과 필연적으로 진화가 일어나게 되는 것임.
이제 본격적으로 이 법칙에 대해 조금 더 알아보자.
p를 대립유전자 A의 빈도라고, q를 대립유전자 a의 빈도라고 해보자. 이 때 gene pool 내에 p, q가 전부라는 가정 하에 p+q =1임.
한편 부모가 자손을 낳았을 시 AA, Aa, aa 중 하나의 유전형을 가지는 자손을 얻을 것임. 이 때 각 유전형에 따른 빈도는 p^2+2pq+q^2으로 나타낼 수 있음. 그런데 이는 곧 (p+q)^2과 같고, 앞서 p+q=1임을 보였으므로 이 값 또한 1이 됨. 즉, 이를 통해 gene pool 내에서의 대립 유전자 빈도는 시간에 따라 변화하지 않는다는 것을 알 수 있음.
다만 Hardy-Weinberg law가 성립하기 위해서는 위 그림에 나타나 있는 5가지 가정이 충족되어야 함. 하나하나 살펴보자.
첫 번째 가정은 selection 없이 동일한 rate로 survival하고 reproduction이 이루어진다는 것임. 그러나 실제로는 natural selection이 작용하므로 이 가정이 성립하지 않음. (참고로 사회적 동물인 인간의 경우 다소 natural selection pressure가 약한 편이어서 이 가정이 어느 정도 들어맞을수도 있음)
두 번째 가정은 새로운 allele이 mutation등에 의해 생기지 않는다는 것임. 그러나 실제로는 DNA replication 과정에서, UV에 노출될 때, chemical 등에 노출될 때와 같이 많은 상황에서 mutation이 발생하므로 이 가정이 성립하지 않음.
세 번째 가정은 population 내로의, 혹은 population 바깥으로의 migration이 일어나지 않는다는 것임. 그러나 실제로는 migration이 많이 일어나므로(초식동물은 풀을 다 뜯어먹고 나면 migration하게 되고, 육식동물은 migration하는 초식동물을 따라 같이 migration하게 됨) 이 가정이 성립하지 않음.
네 번째 가정은 population의 크기가 무한하다는 것임. 그러나 실제로는 population의 크기가 유한하므로 이 가정이 성립하지 않음.
마지막 가정은 mating이 random하게 일어난다는 것임. 이 가정의 경우 성립되는 경우도 있고 성립되지 않는 경우도 있음. 성립되지 않는 경우를 예를 들어 생각해보면, 특정 생물이 자신 주변의 작은 pool 내에서만 주로 교배할 경우 이는 완전한 random mating이 아니게 되므로 이 가정이 깨지게 됨.
Hardy-Weinberg law의 중요한 3가지 결론은 다음과 같음.
1. 우성형질(dominant traits)이 세대를 거칠수록 반드시 더 증가하는 것은 아님.
2. 유전적 다양성은 유지될 수 있음.
3. 단지 한 개의 유전형 빈도만을 알아도 다른 유전형의 빈도들을 계산해낼 수 있음.
이제 Hardy-Weinberg law를 인간 집단에 대해 적용해보자.
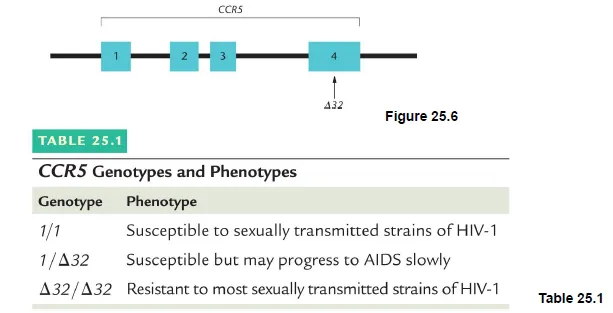
예를 들어 HIV-1 virus 감염의 감수성에 영향을 미치는 유전자에 대해 생각해보자. 실제로 위 그림에 나타나 있는 것처럼 CCR5 gene의 homozygous Δ32(mutant)를 가지는 경우 HIV-1에 잘 감염되지 않음. 그 이유는 CCR5가 T cell의 receptor여서 원래는 HIV virus가 감염될 시 이 receptor에 의해 인지되게 되는데, Δ32 homozygote의 경우 HIV virus가 T cell에 의해 인식되지 못하게 되므로 그 결과 infection이 잘 일어나지 않게 되는 것임.
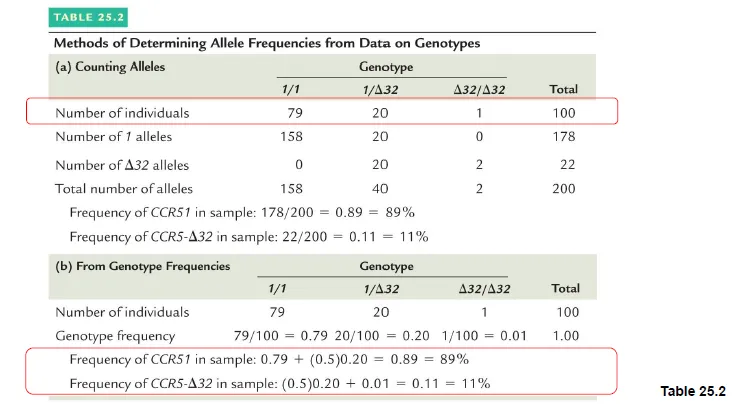
실제로 위 그림 (a)에서와 같이 allele의 수가 counting되어 주어졌다고 해보자. 이 경우 CCR5-1의 frequency는 {158+40/2}/{200} x 100 (%) = 89%로 계산됨. 한편 CCR5- Δ32의 frequency는 {2+40/2}/{200} x 100 (%) = 11%로 계산됨.
이 밖에 위 그림 (b)에서와 같이 주어진 genotype frequency를 이용해서도 각 allele의 frequency를 계산할 수 있으므로 참고할 것.
한편 지금까지의 논의와 유사하게, 다수의 유전자에 대해서도 (p+q+r+...)^2과 같은 식의 표현법을 통한 계산을 수행할 수 있음. 참고로 그 밖에, X 염색체 연관 형질의 대립유전자 빈도를 계산할 떄는 gene pool에서 X 염색체와 관련된 대립유전자의 빈도가 남성이 X 염색체 연관 형질을 발현하는 빈도와 동일하다는 것을 기억해두면 됨. (왜냐하면 남자는 X 염색체가 한개 뿐이기 때문)
한편 Hardy-Weinberg law를 이용해 이형접합자의 빈도도 계산할 수 있음. 예를 들어 어떤 집단에서 cystic fibrosis의 발병률이 0.0004라고 해보자. 이 때 이와 관련된 recessive allele의 빈도를 q라고 하면 q는 Hardy-Weinberg law에 따라 아래와 같이 계산될 수 있음.
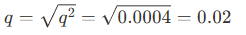
한편 Hardy-Weinberg law에 따라 p+q = 1이므로 p =0.98이 됨. 그러므로 이형접합자의 빈도는 2pq = 0.04로 계산될 수 있음.
다음 포스트부터는 Hardy-Weinberg law가 성립하지 않는 실제 상황에 대해 조금 더 논해보자.
'전공자를 위한 생물학 > 유전학' 카테고리의 다른 글
| [유전학] 12.3 : 대진화(macroevolution) (0) | 2024.11.02 |
|---|---|
| [유전학] 12.2 : 하디 바인베르크 법칙의 예외 (0) | 2024.11.02 |
| [유전학] 11.3 : Hox 유전자 발현 조절 (0) | 2024.11.02 |
| [유전학] 11.2 : 척추동물에서의 Hox 유전자 (0) | 2024.11.02 |
| [유전학] 11.1 : 초파리의 발생과 Hox 유전자 (0) | 2024.11.02 |