이전 포스트까지는 원핵생물이 operon이라는 system을 이용해서 제한적인 숫자의 gene transcription을 조절하는 것을 보았음.
이번 포스트부터는 원핵생물에서 일어나는 보다 큰 규모의 유전자 발현 조절 기작들에 대해 알아보자.
크게 아래와 같은 3가지 메커니즘이 대표적임.
1) sigma factor switching
2) RNA polymerase switching
3) antitermination
이 중에서 이번 포스트에서는 특별히 sigma factor switching에 대해 알아보자.
sigma factor switching은 일반적으로 bacteria에 phage가 infection되는 경우 일어나며, 이 때 침입한 phage의 gene이 temporal하게 발현하므로 이 과정에서 단계적으로 sigma factor switching이 일어남.
흔히 sigma factor switching을 이용하는 phage는 T4와 SPO1 등이 있음. (이 둘 모두 꽤나 큰 genome을 가지고 있음) 그 중에서 특히 B. subtilis에 감염되는 SPO1 phage에 대한 연구가 많이 이루어졌으므로 이에 대해 살펴보자.

위 그림에서 early transcription stage는 phage가 감염되고 난 후 5분동안에 해당하며 이 시기에 early gene의 발현이 일어남. 이 때는 (a)에서도 알 수 있듯 bacteria의 core protein, sigma factor를 그대로 사용해 gene expression이 일어남. 다음으로 감염 후 5~10분동안은 middle gene의 발현이 일어남. 이 때는 (b)에서도 알 수 있듯 bacteria의 core protein과 더불어 (a)에서 SPO1의 gene으로부터 합성된 gp28이 sigma factor로 사용됨. 이렇게 되면 gp28이 middle gene의 promoter만을 특이적으로 인식할 수 있어서 결과적으로 middle gene만 전사됨. 마지막으로 감염 10분 후에는 late gene이 expression됨. 이 때 (c)에서도 알 수 있듯 이 때는 middle gene으로부터 발현된 gp33, gp34가 합쳐져서 앞서와는 완전히 다른 sigma factor를 구성해 bacteria의 core protein과 결합함. 그 결과 late gene이 특이적으로 전사됨.

이와 관련한 실험 결과가 위에 나타나 있음. 보면 SPO1이 감염된 후 시간이 좀 지난 bacteria에서 추출한 단백질을 대상으로 gel electrophoresis를 수행해보았는데, 이 때 sigma subunit에 해당하는 band는 나타나지 않고 그 대신 gp28, gp34, gp33에 해당하는 band가 나타남. 이를 통해 시기별로 gp28, gp34, gp33이 sigma factor의 역할을 수행한다는 것을 알 수 있음.
지금까지 살펴본 SPO1에서의 sigma factor switching 현상은 sporulation(포자형성)에서도 유사하게 관찰됨. B. subtilis와 같은 특정 bacteria는 영양소가 풍부한 환경 하에서는 vegetative state로 존재하지만, 만약 starvation condition에 처하게 되면 이 녀석은 dormant body(endospore)를 형성한 후 favorable condition이 다시 돌아올 때까지 언제까지고 동면하고 있게 됨. 이 때 vegetative state에서 발현되는 gene과 sporulation 시 발현되는 gene의 종류는 완전히 다르며, 이들의 시기별 turn-on, turn-off 시 특정한 종류의 sigma factor가 관여하게 됨. (vegetative sigma factor와 sporulous sigma factor가 따로 존재하는데, 이들은 각각 서로 잘 결합하는 promoter들이 있게 되고 그 결과 이들 중 어떤 녀석이 있느냐에 따라 대규모의 전사조절이 일어나게 됨)
한편 sporulation, vegetative state 모두에서 turn-on되어 있어야만 하는 gene도 있을 것임. 그렇다면 이들은 어떻게 해서 sigma factor가 바뀜에도 불구하고 계속 turn-on되어 있을 수 있을까.
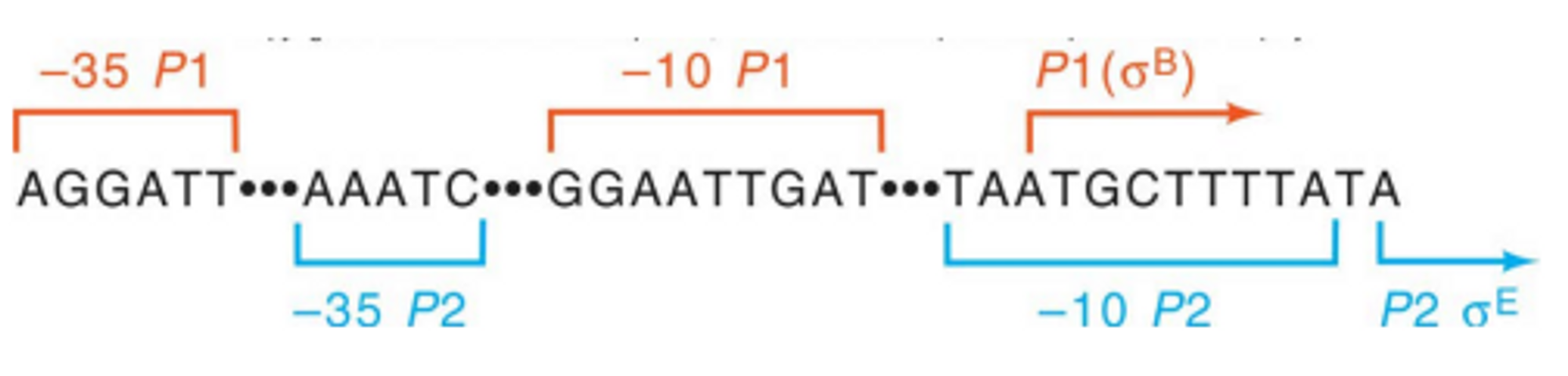
이들은 위 그림과 같이 두 개 이상의 promoter를 가지고 있음. 따라서 sigma factor가 바뀌더라도 이 gene은 (10bp가량의 일부 겹치지 않는 영역을 제외하고) 잘 발현될 것임.

위 그림은 앞서 설명한 multiple promoter를 가지는 gene에 대한 실험 결과임. 이 때 sigmaE의 존재하에서는 110nt길이의 RNA가, sigmaB의 존재하에서는 120nt 길이의 RNA가, sigmaE, sigmaB가 둘 다 존재할 때에는 110, 120nt 길이의 RNA 모두가 관찰됨을 확인할 수 있음. 이를 통해 (비록 10nt정도의 길이차이는 있지만) sigma factor가 달라져도 multiple promoter를 가지고 있으므로 계속 발현되는 gene이 존재한다는 것을 알 수 있음.
한편 bacterial heat shock 상황에서 heat shock response가 일어나는데, 이 때 이 response는 온도가 급격하게 높아짐에 따라 생길 수 있는 damage를 최소화하기 위한 기작임. heat shock response의 가장 눈에 띄는 결과 중 하나는 molecular chaperone을 더 많이 발현시키거나, 그 밖의 heat shock gene들을 더 많이 발현시키는 것임. 이 때 평소에는 꺼져있던 이들 gene의 발현을 turn-on 시키는 데에도 지금까지 배워온 것과 같은 sigma factor switching이 이용됨.
특히 E. coli에서 heat shock response시 사용되는 특정한 sigma factor는 sigma32, sigmaH인데(이 때 H는 heat shock을 의미) 이들은 heat shock gene promoter에 RNA polymerase가 달라붙도록 해줌. (이 때 사실 sigma32, sigmaH 이외에 기존에 사용되던 sigma factor도 존재할 것이므로 결국 sigma32, sigmaH와 기존의 sigma factor가 서로 core protein과 결합하는 것에 대한 일종의 경쟁을 펼치게 될 것임) 그렇다면 높은 온도 환경에서 sigmaH가 많아질 수 있는 이유는 무엇일까. 크게 아래와 같은 두 가지 상황이 가능함.
1) 온도 상승에 따라 sigmaH의 구조가 안정화됨.
2) 온도가 높아짐에 따라 sigmaH를 encoding하는 mRNA의 translation이 촉진됨.
이 밖에 low nitrogen 혹은 starvation stress 등에 의해 유발되는 response들도 마찬가지로 sigma factor switching에 의해 특정 유전자를 특이적으로 turn-on 시키는 기작을 사용함.
한편 anti-sigma factor라는 것도 존재함. 이 녀석은 특정 sigma factor를 잡아서 이 sigma factor가 core protein과 작용하지 못하도록 막아주는 역할을 함. (따라서 평소에는 이 anti-sigma factor가 sigma factor를 억제하고 있다가, 특정 event가 발생했을 때애만 anti-sigma factor가 불활성화, 혹은 분해되는 식으로 temporal한 sigma factor switching의 조절에 도움을 줄 수 있음) 이에 대한 대표적인 예가 E. coli가 가지는 rsd gene으로부터 만들어지는 단백질임.
이 밖에 anti-sigma factor와 sigma factor가 형성한 complex에 결합해 anti-sigma factor를 release시켜주는 역할을 하는 anti-anti-sigma factor도 존재함.
다음 포스트에서는 RNA polymerase switching에 의한 유전자 발현 조절기작에 대해 알아보자.
'전공자를 위한 생물학 > 분자생물학' 카테고리의 다른 글
| [분자생물학] 8.3 : lambda phage(람다 파지), antitermination - 1 (0) | 2023.12.16 |
|---|---|
| [분자생물학] 8.2 : RNA polymerase switching (0) | 2023.12.16 |
| [분자생물학] 7.3 : trp operon(tryptophan operon, 트립토판 오페론), riboswitch(리보스위치) (0) | 2023.08.31 |
| [분자생물학] 7.2 : ara operon(arabinose operon, 아라비노스 오페론) (0) | 2023.08.31 |
| [분자생물학] 7.1 : lac operon(젖당 오페론) - 4 (0) | 2023.08.31 |