[생화학] 18.3 : 산화적 인산화를 통한 ATP 합성 - 3
이전 포스트에 이어서 살펴보자.
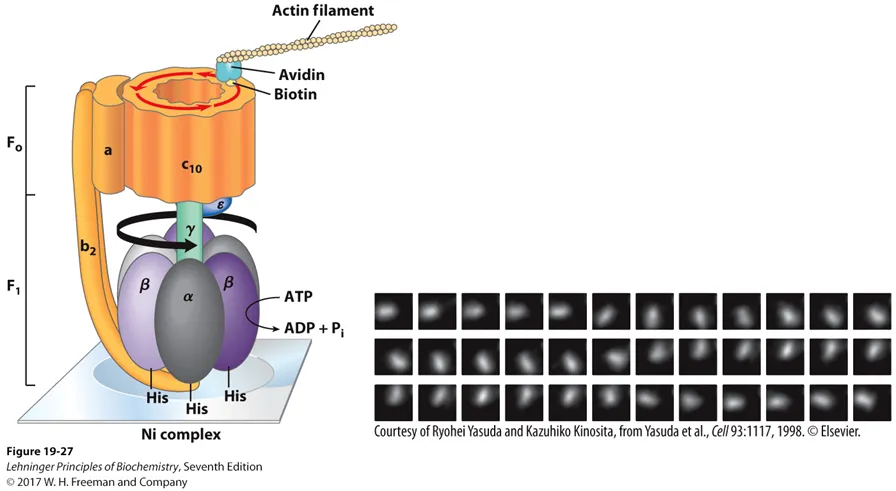
앞서 계속 F1의 상대적인 position에 대해 이야기했었는데, 사실 이 position은 위 그림상에서 central shaft γ에 대한 상대적인 position임. 실제로 proton이 F0를 통과하면서 F0의 rotation이 일어나게 되면 \gamma shaft도 rotation되게 됨. 한편 F1은 고정되어 있게 되고 결과적으로 \gamma shaft에 대한 F1 내의 dimer 각각의 위치가 달라지게 됨. (이런 일이 발생할 수 있는 것은 어디까지나 γ shaft가 비대칭적이어서 회전에 따른 차이를 만들어낼 수 있기 때문임)
위 그림의 경우 특별히 F0 중 직접적인 회전이 일어나는 부위인 C10(C subunit 10개로 이루어져 있음)이 정말로 회전하는지를 확인하기 위해 이곳에 biotin-avidin tagging을 이용해 actin filament tag를 붙인 후 electron microscopy로 관찰한 결과를 보여주고 있음. 실제로 위 그림 오른쪽 data를 보면 C10이 360도 회전한다는 것을 알 수 있음.
그렇다면 대체 어떤 기작에 의해 proton이 통과하는 과정에서 F0의 rotation이 일어나게 되는 것일까.
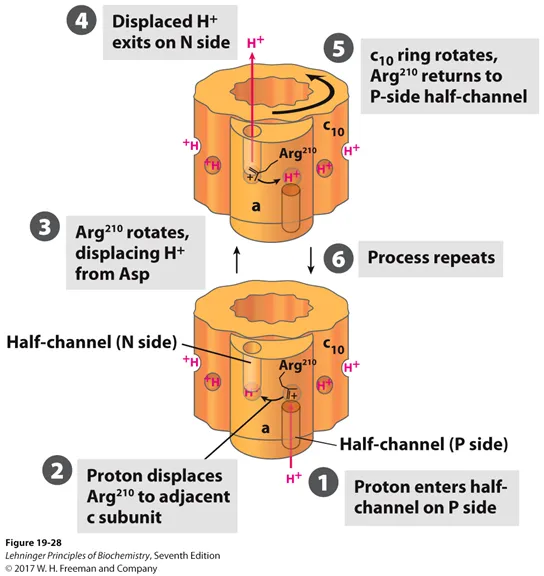
이와 관련된 모식도가 위 그림상에 나타나 있음. 하나하나 살펴보자.
위 그림을 보면 a subunit이 그림상에서 앞쪽에 표시되어 있으며, 그 뒤에 c subunit이 존재하고 있음. 이 때 a subunit에 H+가 통과할 수 있는 channel이 있는데, 이 channel이 그냥 통으로 뚫려있는게 아니라 2개의 half-channel로 split되어있음. 그러다보니 proton이 바로 이곳을 통해 통과하지는 못하고 통과 과정에서 c subunit의 rotation을 유발하게 되는 것임.
우선 1에서 나타난 것처럼 P side에 있는 half-channel로 proton이 들어옴. 그러면 이 proton은 C subunit에 있는 Asp의 COO-와 정전기적 상호작용을 하며 결합하게 됨. 그럴 시 원래 Asp와 상호작용을 하고 있었던 a subunit에 있는 Arg가 상호작용을 더이상 하지 못하게 되고 Arg residue가 3과 같이 옆칸으로 kick out되어버림. 그럴 시 이렇게 쫒겨난 Arg는 그 자리 근방의 Arg와 상호작용을 하게 되고, 그 결과 원래 상호작용을 하고 있던 H+ ion을 kick out시켜버림. 이렇게 해서 내쫒아진 H+는 N side의 half-channel을 통해 mitochondrial matrix 방향으로 빠져나가버리게 됨. 이후 Arg과 Asp의 interaction에 의해서 위 그림상에서 시계반대방향으로의 (한칸만큼) 회전이 일어나게 되며 처음과 동일한 상황이 됨. (참고로 위 그림상에서 시계 반대방향으로 이동하므로 결국 맨 첫 step에서 들어간 proton은 한바퀴를 다 돌고나야 비로소 N side의 half-channel을 통해 matrix로 빠져나갈 수 있음)
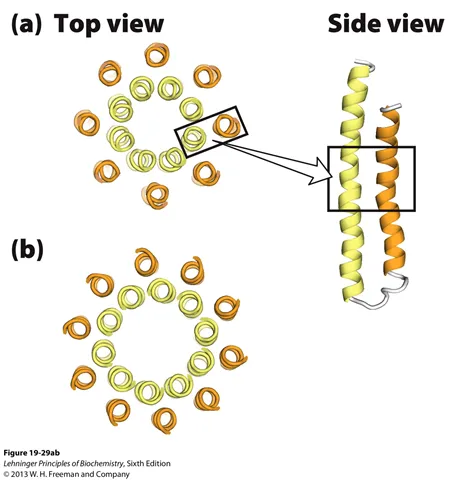
위 그림에는 F0 complex의 C ring 구조가 나타나 있음. 이 때 주황색 부근에 Asp이 존재하고 있음.
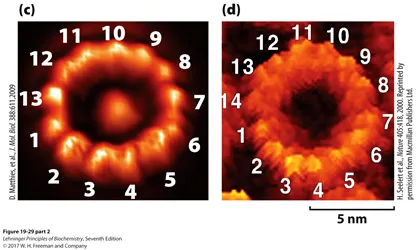
한편 위 그림과 같이 종에 따라서 c ring을 구성하는 c subunit의 갯수가 다름. 그러다 보니 결국 한바퀴를 돌리기 위해 필요한 proton의 수도 달라지는 것임.
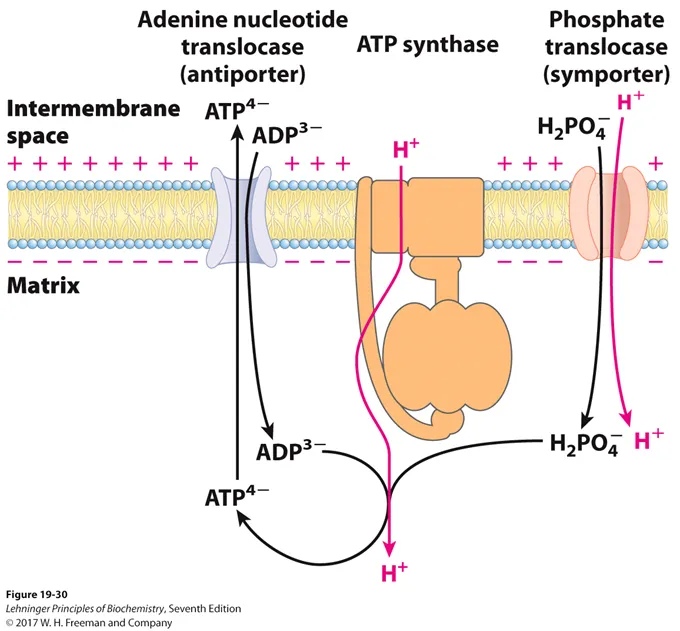
한편 위 그림은 앞서 잠시 언급했었던 ATP, ADP, P_i의 mitochondria 내외로의 수송 과정을 나타내 보여주고 있음. 일단 mitochondria 내막에 있는 adenine nucleotide translocase는 ATP는 밖으로, ADP는 안으로 수송해주는 antiporter로 기능함. (이 때 이 수송은 ATP, ADP의 농도에 dependent하게 일어나므로 별도의 energy가 필요하지 않음) 한편 mitochondria 내부에서는 ATP 합성이 지속적으로 일어나야 하므로 ADP 뿐만 아니라 Pi도 필요한데, 이 Pi를 H2PO4-의 형태로 mitochondria 안으로 수송시켜주는 녀석이 바로 phosphate translocase임. 이 녀석은 H2PO4-와 H+를 같은 방향으로 수송시켜주는 symporter로 기능함. (이 때 phoshpate를 matrix 내부로 수송시키는 과정에서 proton gradient 일부를 소모해야 한다는 것을 알 수 있음)
따라서 많은 과학자들은 ATP가 생성되는 당량을 계산할때 이를 고려해주기도 함. 일반적으로 진핵생물에서 ATP synthase를 통해 ATP 한 분자를 형성시키기 위해 필요한 proton 갯수는 대략 3개로 보며, 이에 더해 방금 봤던 것처럼 phosphate 한 분자를 matrix로 이동시키는데 추가적인 proton 1개가 더 사용되므로 결국 ATP 한 분자 합성을 위해서 proton 4개분의 gradient가 쓰인다는 것을 알 수 있음. 한편 앞서 봤듯이 NADH 한 분자에 의해서는 proton 10개만큼이 pumped out되며, FADH2 한 분자에 의해서는 proton 6개만큼이 pumped out됨. 따라서 NADH 한 분자의 경우 10/4 = 2.5개의 ATP 생산에 기여할 수 있고, FADH2 한 분자의 경우 6/4 = 1.5개의 ATP 생산에 기여할 수 있음.
다음 포스트에서 이어서 살펴보자.